The ubiquitin proteasome system (UPS) is part of the waste management system of a cell. Proteins that are destined to have limited life spans in the cell are identified and sorted for destruction. This is accomplished by labeling the proteins with the molecule ubiquitin, which is present in all eukaryotes. Once tagged, the proteins are degraded by a complex machine known as the proteasome. The UPS is not just a molecular wood chipper; it performs its function with specificity that is provided by the class of ubiquitination enzymes called E3 ligases. The E3 ligases identify proteins to be degraded through domains called F-boxes, but the mechanism of enzymatic transfer of ubiquitin to the doomed protein is still not completely understood and may be facilitated by the dimerization of another domain, known as the D-domain. In studies carried out at the NE-CAT 8-BM-B, BioCars 14-BM-C, BioCAT 18-ID-D, and SBC-CAT 19-BM-D beamlines, all at the APS, researchers solved the structures of two of these D-domains: one from the yeast protein Cdc4 (2.5 Å) and another from human β-TrCP (2.4 Å). This work provides new insights into the mechanism of the E3 ligases by revealing that D-domain dimerization plays a role in positioning different substrates for ubiquitination.
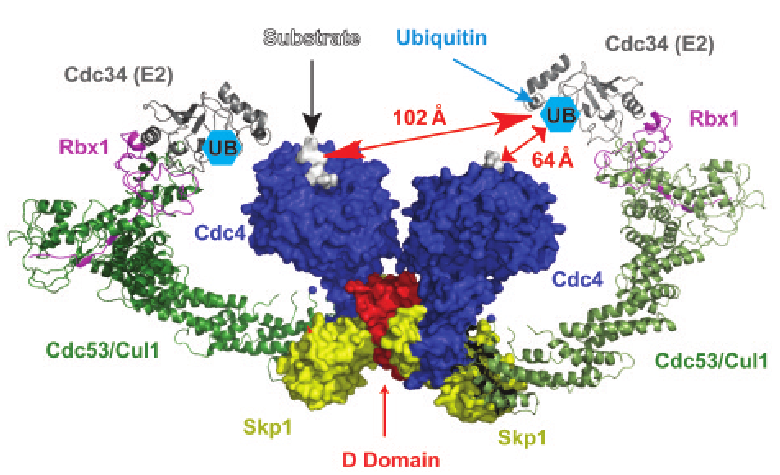
Using a combination of x-ray crystallography, mutational analysis, and small-angle x-ray scattering (SAXS) measurements, the researchers from the Samuel Lunenfeld Research Institute, the Cellgene Corporation, the University of Western Ontario, and the University of Toronto were able to reveal important new information regarding the role of the D-domain in the E3 ligase. Solution of the structures of the Cdc4 (Fig. 1) and β-TrCP D-domains reveals a common fold. Each fold is composed of three α helices that associate in a right-handed superhelical manner to form a parallel dimer. Of interest to the function of these proteins is that the sequences of the D-domains for Cdc4 and β-TrCP share only two conserved residues among the 16 amino acids involved in the dimerization interface. However, in both structures the dimerization interface is hydrophobic in nature, with variability between the two proteins imparting specificity of binding between different D-domains.
In order to learn more about how D-domain dimerization affects E3 ligase functions such as substrate recognition and ubiquitin conjugation, the team performed mutational analyses on the Cdc4 molecule. They found that dimerization of the D-domain is essential for the function of the full-length Cdc4 protein and showed that two residues were critical for dimer formation. However, although Cdc4 proteins that contained mutated D-domains did not dimerize, they were still able to bind substrates. Analysis of the catalytic activity of the ubiquitination reaction, however, revealed that the catalytic attachment of ubiquitin was impaired in these mutants. Therefore, it appears that dimerization of Cdc4 mediated through the D-domain is important for catalytic efficiency but not for substrate recognition.
SAXS analysis provided further insight into the importance of D-domain dimerization and its involvement in the catalytic efficiency of the ubiquitination reaction. Catalytic attachment of multiple ubiquitin molecules to lysine residues in a protein that may be spaced at different distances along the protein presents a situation that would be facilitated by an accommodating catalytic site. SAXS analysis of Ddomain plus F-box domain constructs revealed that, although the distance between these domains is quite constrained, the configuration created by dimerization brings the substrate binding domain of the F-box into coplanar contact with the E2 catalytic enzyme. This allows for a wide variety of geometric configurations to fit within the catalytic domain, increasing catalytic efficiency. Together, this analysis of Ddomain dimerization suggests that the flexibility of the system comes from dimerization imposed by the D-domain as shown in the crystal structure and the coplanar configuration created by dimerization that allows substrates of varying sizes and configurations to be available for polyubiquitination and subsequent disposal by the proteasome.
— Sandy Field
See: Xiaojing Tang, Stephen Orlicky, Zhenyuan Lin, Andrew Willems, Dante Neculai, Derek Ceccarelli, Frank Mercurio, Brian H. Shilton, Frank Sicheri, and Mike Tyers, “Suprafacial Orientation of the SCFCdc4 Dimer Accommodates Multiple Geometries for Substrate Ubiquitination,” Cell 129, 1165 (June 15, 2007). DOI 10.1016/j.cell.2007.04.042
This work was supported by grants to B.H.S., F.S., and M.T. from the Canadian Institutes of Health Research, the National Cancer Institute of Canada (NCIC), and the Natural Sciences and Engineering Research Council. BioCAT is National Institutes of Health-supported Research Center RR-08630. Use of the Advanced Photon Source was supported by the U.S. Department of Energy, Office of Science, Office of Basic Energy Sciences, under Contract No. DE-AC02-06CH11357.