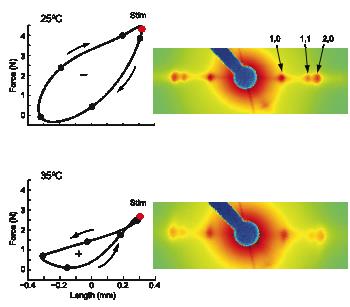
The Hawkmoth Manduca sexta is an emerging model system for a wide range of studies in integrative biology. The flight muscles are particularly interesting in that, unlike most insect flight muscle, but like vertebrate skeletal and cardiac muscles, they are a synchronous muscle where each stimulus generates one muscle twitch. The length tension curve also shows intriguing similarities to mammalian cardiac muscle even though the sarcomere structure is known to be quite different. Another property of the muscle is that the dorsal-most region of the flight muscle is ca. 5 degrees C cooler than the ventral muscle closer to the midline to the body. (Such spatial temperature gradients are also likely to occur in large muscles in mammals but this has not been well investigated). In Manduca flight muscle these spatial gradients lead to a spatial variation in power production spanning from positive to negative values across the predicted temperature range. Warm ventral subunits produce positive power at their in vivo operating temperatures, and therefore act as motors producing force. Concurrently, as muscle temperature decreases dorsally, the subunits produce approximately zero mechanical power output. These muscles, therefore, not only generate force, but also may act as springs, providing energy storage to drive locomotion. The question addressed in this study is what component of the muscle enables the colder muscle to act as springs.
Using time-resolved small-angle X-ray diffraction paired with in situ measurements of mechanical energy exchange in the flight muscle of Manduca sexta the authors produced 5-frame, 8 ms per frame, movies of the X-ray equatorial reflections indicating cross-bridge association with myofilaments. These movies showed cyclical changes in the intensities of the equatorial reflections in the warmer, force generating muscle and stable equatorial intensities in cooler muscles acting as springs. These results indicate that the temperature gradient creates a locked-spring lattice in the cooler region of muscle. Cross-bridges in that region that remain bound and elastically deformed at the end of muscle shortening could release the stored strain energy during muscle lengthening. These results suggest that cross-bridges can perform functions other than just contraction, including acting as molecular links for elastic energy storage.
See: N.T. George, T.C. Irving, C.D. Williams, and T.L. Daniel “ The cross-bridge spring: cool muscles store elastic energy” Science 340, 1217-1220 (2013)